
Emergence of Maps in the Memories of Blind Navigation Agents
Introduction
Decades of research into intelligent animal navigation posits that organisms build and maintain inter- nal spatial representations (or maps) of their environment, that enables the organism to determine and follow task-appropriate paths. Hamsters, wolves, chimpanzees, and bats leverage prior exploration to determine and follow short- cuts they may never have taken before. Even blind mole rats and animals rendered situationally-blind in dark environments demonstrate shortcut behaviors. Ants forage for food along meandering paths but take near-optimal return trips, though there is some controversy about whether insects like ants and bees are capable of forming maps.
Analogously, mapping and localization techniques have long played a central role in enabling non- biological navigation agents (or robots) to exhibit intelligent behavior. More recently, the machine learning community has produced a surprising phenomenon – neural-network models for navigation that curiously do not contain any explicit mapping modules but still achieve remarkably high performance For instance, Wijmans et al. (2020) showed that a simple ‘pixels-to-actions’ architecture (using a CNN and RNN) can navigate to a given point in a novel environment with near-perfect accuracy; Partsey et al. (2022) further generalized this result to more realistic sensors and actuators. Reed et al. (2022) showed a similar general purpose architecture (a transformer) can perform a wide variety of embodied tasks, including navigation. The mechanisms explaining this ability remain unknown. Understanding them is both of scientific and practical importance due to safety considerations involved with deploying such systems.
In this work, we investigate the following question – is mapping an emergent phenomenon? Specifically, do artificial intelligence (AI) agents learn to build internal spatial representations (or ‘mental’ maps) of their environment as a natural consequence of learning to navigate?
Task
The specific task we study is PointGoal navigation, where an AI agent is introduced into a new (unexplored) environment and tasked with navigating to a relative location – ‘go 5m north, 2m west relative to start’. This is analogous to the direction and distance of foraging locations communicated by the waggle dance of honey bees.
Agent input and design
Unlike animal navigation studies, experiments with AI agents allow us to precisely isolate map- ping from alternative mechanisms proposed for animal navigation – the use of visual land- marks orientation by the arrangement of stars, gradients of olfaction or other senses. We achieve this isolation by judiciously designing the agent’s perceptual system and the learning paradigm such that these alternative mechanisms are rendered implausible. Our agents are effectively ‘blind’; they possess a minimal perceptual system capable of sensing only egomotion, i.e. change in the agent’s location and orientation as the it moves – no vision, no audio, no olfactory, no haptic, no magnetic, or any other sensing of any kind. This perceptual system is deliberately impoverished to isolate the contribution of memory, and is inspired by blind mole rats, who perform localization via path integration and use the Earth’s magnetic field as a compass. Further still, our agents are composed of navigation-agnostic, generic, and ubiquitous architectural components (fully-connected layers and LSTM-based recurrent neural networks), and our experimental setup provides no inductive bias towards mapping – no map-like or spatial structural components in the agent, no mapping supervision, no auxiliary tasks, nothing other than a reward for making progress towards a goal.
Results
Surprisingly, even under these deliberately harsh conditions, we find the emergence of map-like spatial representations in the agent’s non-spatial unstructured memory, enabling it to not only successfully navigate to the goal but also exhibit intelligent behavior (like taking shortcuts, following walls, detecting collisions) similar to aforementioned animal studies, and predict free-space in the environment. Essentially, we demonstrate an ‘existence proof’ or an ontogenetic developmental ac- count for the emergence of mapping without any previous predisposition. Our results also explain the aforementioned surprising finding in recent literature – that ostensibly map-free neural-network achieve strong autonomous navigation performance – by demonstrating that these ‘map-free’ systems in fact learn to construct and maintain map-like representations of their environment.
Concretely, we ask and answer following questions:
Is it possible to effectively navigate with just egomotion sensing?
Yes. We find that our ‘blind’ agents are highly effective in navigating new environments – reaching the goal with 95.1%$\pm$1.3% success rate. And they traverse moderately efficient (though far from optimal) paths, reaching 62.9%$\pm$1.6% of optimal path efficiency. We stress that these are novel testing environments, the agent has not memorized paths within a training environment but has learned efficient navigation strategies that generalize to novel environments, such as emergent wall-following behavior.
What mechanism explains this strong performance by ‘blind’ agents?
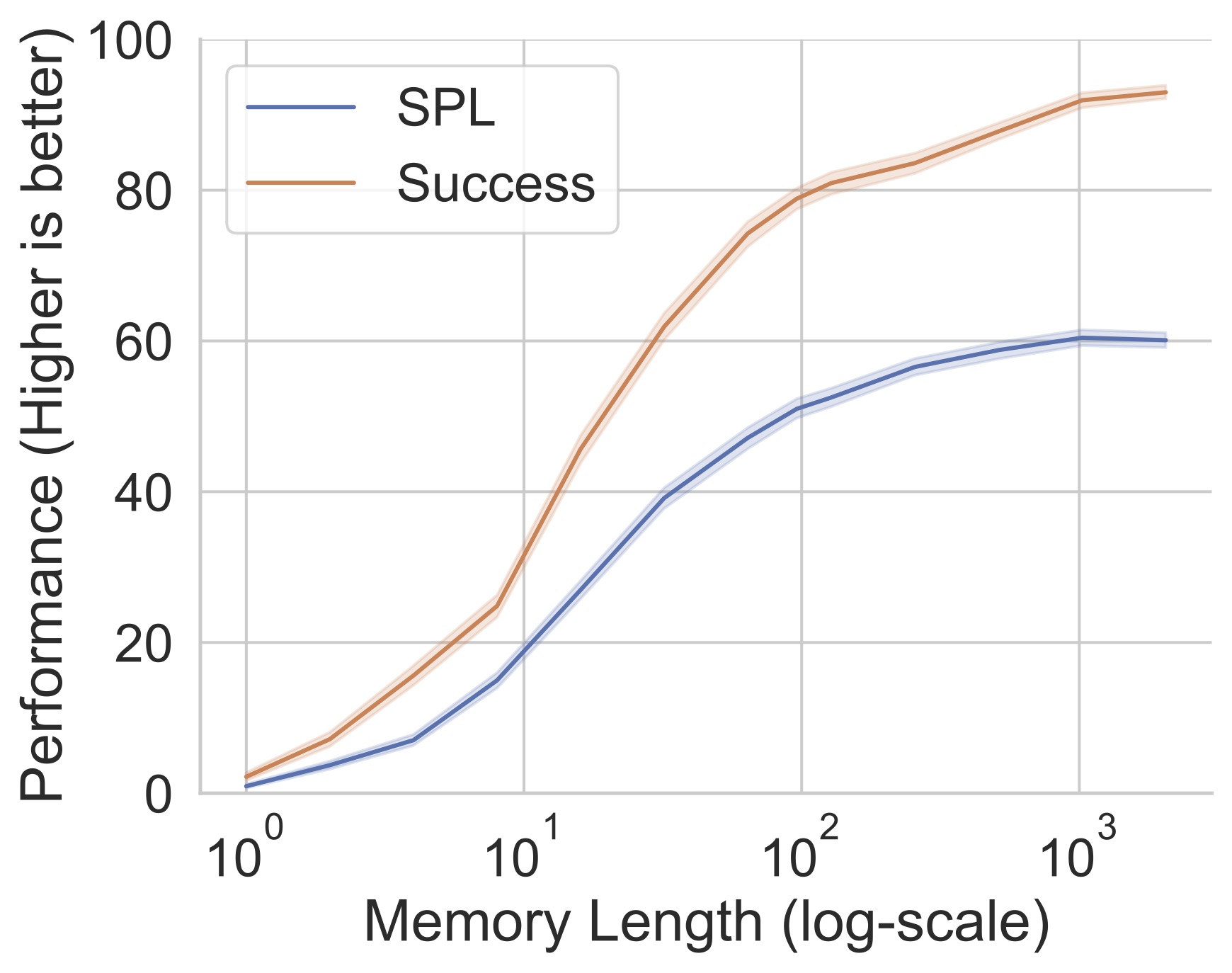
Memory. We find that memoryless agents completely fail at this task, achieving nearly 0% success. More importantly, we find that agents with memory utilize information stored over a long temporal and spatial horizon and that collision-detection neurons emerge within this memory. Navigation performance as a function of the number of past actions/observations encoded in the agent’s memory does not saturate till one thousand steps (corresponding to the agent traversing 89.1$\pm$0.66 meters), suggest- ing that the agent ‘remembers’ a long history of the episode.
The video bellow shows a video of a blind agent navigating (right) and the t-SNE projection of the portion of it’s hidden state responsible for detecting collision. The black dot is where the agent’s hidden state is for the current state. Notice that the hidden state stays within the same cluster throughout a series of actions.
What information does the memory encode about the environment?
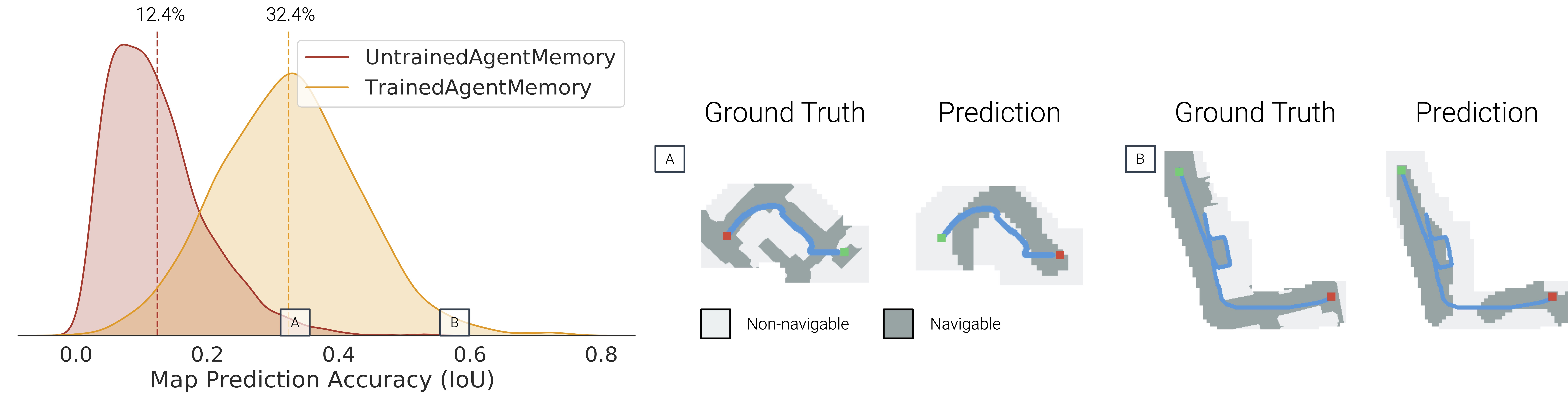
Implicit maps. We perform an AI rendition of Menzel (1973)’s experiments, where a chimpanzee is carried by a human and shown the location of food hidden in the environment. When the animal is set free to collect the food, it does not retrace the demonstrator’s steps but takes shortcuts to collect the food faster. Analogously, we train a blind agent to navigate from a source location (S) to a target location (T). After it has finished navigating, we transplant its constructed episodic memory into a second ‘probe’-agent (which is also blind). We find that this implanted-memory probe-agent performs dramatically better in navigating from S to T (and T to S) than it would without the memory transplant. Similar to the chimpanzee, the probe agent takes shortcuts, typically cutting out backtracks or excursions that the memory-creator had undertaken as it tried to work its way around the obstacles. These experiments provide compelling evidence that blind agents learn to build and use implicit map-like representations of their environment solely through learning to navigate. Intriguingly further still, we find that surprisingly detailed metric occupancy maps of the environment (indicating free-space) can be explicitly decoded from the agent’s memory.
This videos show a blind agent navigating from source to target and then a probe navigating from target to source. Notice the increase efficiency and reduction in collisions of the probe during the return trip.
Are maps task-dependent?
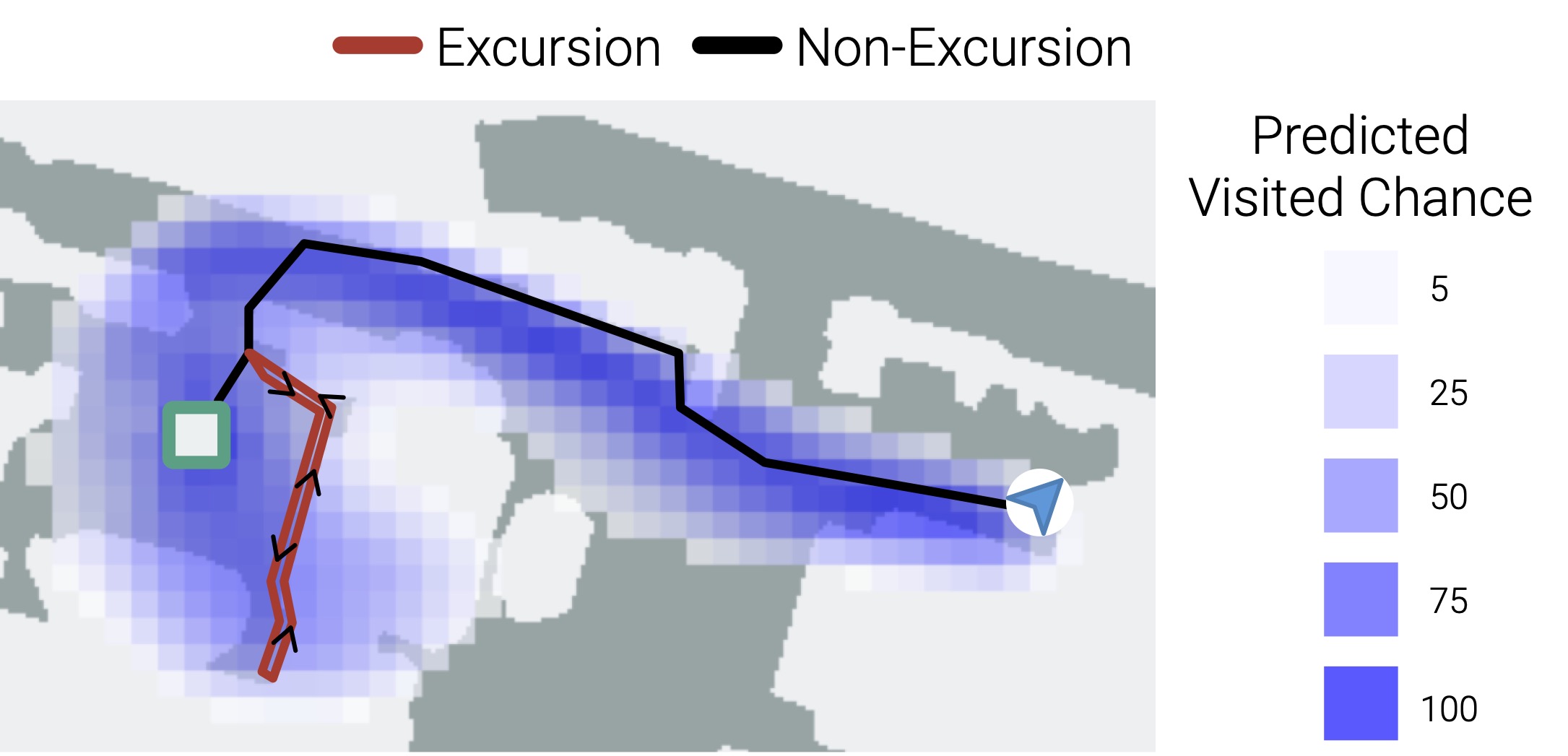
Yes. We find that the emergent maps are a function of the navigation goal. Agents ‘forget’ excursions and detours, i.e. their episodic memory only preserves the features of the environment relevant to navigating to their goal. This, in part, explains why transplanting episodic memory from one agent to another leads it to take shortcuts – because the excursion and detours are simply forgotten.
Conclusion
Overall, our experiments and analyses demonstrate that ‘blind’ agents solve PointGoalNav by combining information over long time horizons to build detailed maps of their environment, solely through the learning signals imposed by goal-driven navigation. In biological systems, convergent evolution of analogous structures that cannot be attributed to a common ancestor (e.g. eyes in vertebrates and jellyfish) is often an indicator that the structure is a natural response to the ecological niche and selection pressures. Analogously, our results suggest that mapping may be a natural solution to the problem of navigation by intelligent embodied agents, whether they be biological or artificial. We now describe our findings for each question in detail.